Modern Times
As dinosaur paleontology entered the 1990s its evolution followed parallel opposing courses. Most dinosaurologists saw birds as dinosaurs, with many of them seeing the latter has having high metabolic rates. An ardent minority denied the dinosaur-bird link and they – plus some who agreed that birds are dinosaurs – denied that the latter were tachymetabolic endotherms. This lead to the hypothesis of gigantothermy, which proposed that the metabolic rates and/or thermoregulation of giant animals converges towards a common level. Based in part on erroneous data that seemed to show that some sea turtles have high metabolisms, there is no doubt that the power production of big reptiles and gigantic filter-feeding sharks is a small fraction of that of mammalian giants. I countered with the hypothesis of terramegathermy, which proposes that land animals over about a tonne must have elevated aerobic power production in order to survive in 1 G. That explains why the largest bradymetabolic land reptiles have never exceeded a tonne. I have also been working to redefine the terminology used to describe the metabolic and thermoregulatory systems of organisms.
Then there was John Ruben. A physiologist, he entered paleo with the intention of bringing painstaking analysis to the field he saw (with some justification) as lacking sufficient scientific rigor. Ruben had done some good work, including helping devise the aerobic capacity hypothesis for the evolution of high metabolics rates. The problem was that he failed to follow his own principles when it came to Mesozoic archosaurs. For reasons that are obscure Science elevated his profile by publishing a series of his studies that have not borne up over time. I found him to be as much politician as scientist. Once, at SVP, I entered the ready room to prep for my upcoming talk. Already there with his team, Ruben apparently thought I was spying – like paleoconferences involved espionage – and demanded I leave the room that was open to every meeting participant. I did not comply. Later I noted online that, contrary to Ruben’s thesis that dinosaur noses were too small to contain water and heat conserving respiratory turbinates, one could park a Buick in the nasal cavity of ceratopsids. This really set Ruben off, he called and warned that further such attacks would result in his becoming my “enemy.”

Which was fine with me. Although it can be annoying, I often enjoy intellectual battles. Of course I make my own mistakes, but exposing the flaws in others' research is fun. As for Ruben, he has largely left the field. I’m still in it. Not surprising considering how the evidence is piling up.
Revolution Resolution
In 1996 the long running dinowar initiated by the Dinosaur Renaissance began to come to a resolution. The source of critical data was the Yixian beds of China, north east of Beijing. As the Chinese economy went free market, local farmers began to illegally excavate and sell fossils from Early Cretaceous lake beds that among other items included hundreds of complete specimens of archaic birds, plus the occasional small theropod bearing adorned with simple feathers, first seen in photos of the type Sinosauropteryx circulated at the New York SVP meeting that year. Ostrom was proud, most were amazed and many were startled. I was neither, having been applying feather insulation to my little theropods for years. The years of biding my time were over. The defining evidence had finally come in. Both high metabolic rates and bird relationships were reinforced.
Some of Bakker’s evidence for dinosaur energetics, such as predator/prey ratios, have fallen by the wayside. And as I have explained in my papers on the subject, dinosaur energetics was variable within the group, as it is within birds and especially mammals whose thermophysiology is so diverse that some are nearly reptilian. The first dinosaurs probably had an extinct, intermediate system. But dinosaurs that lived in polar winter habitats too cold and snowy for reptiles, the high walking speeds recorded in their trackways, the high circulatory pressures demanded by their often high held heads, bone isotope analysis, small theropod skeletons found in incubating posture atop their eggs, feathers, and the rapid growth indicated by bone histology have furthered the growing consensus that dinosaurs great and small had higher metabolic rates and aerobic exercise capacity than reptiles.
 It was time to present my work on bird origins, with emphasis on the neoflightless hypothesis, in a technical book. Contacted the science editor at Johns Hopkins Press, Ginger Berman, and made the arrangements. In 2002 Dinosaurs of the Air was released. It was, ironically, already out of date, things move so fast these days. The neoflightless hypothesis predicts that basal dromaeosaurs had fully developed wings even better adapted for flight than Archaeopteryx. I already suspected that was true of the Yixian Sinornithosaurus, whose central finger was flattened bird-style to better support an outer wing. But the feathers were not preserved. Then came along its very close and smaller relative, which records well-developed wings on both the arms and – here even I was surprised – the legs. Way cool. The view that flightless dromaeosaurs were secondarily flightless is now widely accepted. If they were not, then dromaeosaurs independently evolved flight and took it to a level beyond that seen in Archaeopteryx.
It was time to present my work on bird origins, with emphasis on the neoflightless hypothesis, in a technical book. Contacted the science editor at Johns Hopkins Press, Ginger Berman, and made the arrangements. In 2002 Dinosaurs of the Air was released. It was, ironically, already out of date, things move so fast these days. The neoflightless hypothesis predicts that basal dromaeosaurs had fully developed wings even better adapted for flight than Archaeopteryx. I already suspected that was true of the Yixian Sinornithosaurus, whose central finger was flattened bird-style to better support an outer wing. But the feathers were not preserved. Then came along its very close and smaller relative, which records well-developed wings on both the arms and – here even I was surprised – the legs. Way cool. The view that flightless dromaeosaurs were secondarily flightless is now widely accepted. If they were not, then dromaeosaurs independently evolved flight and took it to a level beyond that seen in Archaeopteryx.
I like alternate history scenarios. The definitive evidence produced by the Yixian dinosaurs leads to an interesting speculation of how things could have gone very differently. In the 1920s the American Museum Chinese expeditions initiated by Osborn hoping to find the ancestors of humans and led by Roy Andrews, headed northwest from Beijing towards the Mongolian badlands. But what if they had spent some time checking out the information about Cretaceous beds on the steppes to the northeast? They might have discovered the plethora of birds, and from that the feathered theropods and biplane dromaeosaurs. Dinosaurology would have been revolutionized before the Great Depression, with birds recognized as dinosaurs, and the latter perceived as non-reptilian in energetics and habits. The “new look” of dinosaurs would have been executed by Knight, et. al. I would have grown up reading Colbert’s books about flying, warm-blooded dinosaurs. The dinosaur world would have been even more fascinating for my young self, and the often annoying debates that still grind on would have been long settled. On the other hand, I would not have enjoyed contributing to the renaissance when it did occur.
In July 2001 a big bash was going to be held for Phil Currie's 25th anniversary in paleontology. I drove out, taking the opportunity to visit my father and stepmother in Indiana on the way. I calculated I could make it from there to central North Dakota in a day, departing ~6:30. But to my shock did not pass the Twin Cities until 4:00, and arrived in downtown Bismarck at 9:30. But no rooms were to be found, so I had to continue north towards the border until found a place at midnight. Total was 1117 miles, which I do not recommend. Next day almost did not make it into Canada because their guards suspected I was going to dig dinosaurs for pay (one fellow who flew from Argentina was actually turned back at the airport for the same reason). The surprise party at the Patricia Bar & Grill was well worth the trip.
Skeleton Keys
As my skeletal library piled up I avoided restoring the skeletons of armored ankylosaurs. They are difficult to do because of their bizarre shape, and for a substantial period a lack of good data. Restorations tended to show ankylosaurs with barrel-shaped bodies almost as deep as they were broad. But the hip complexes of ankylosaurs show the ilia flaring far out to the sides to support a fantastically broad ilia, attached to the vertrebral column by extremely long ribs, forming a flat, table-like surface much broader than deep. I finally got around to doing Euoplocephalus, a dinosaur one could set up a picnic spread upon. Have since done a number of other ankylosaurs. The top views usually show the armor on the right side. Can be quite tedious inking out all those little ossicles, one after another.
 In the early 1990s I received a call from Gakken publishers in Japan and began to contribute to their Dino Frontline series. From this evolved The Complete Illustrated Guide to Dinosaur Skeletons in 1996, which like PDW is out of print and I have no particular advice on where to obtain a copy. Sadly, the editor died at a young age.
In the early 1990s I received a call from Gakken publishers in Japan and began to contribute to their Dino Frontline series. From this evolved The Complete Illustrated Guide to Dinosaur Skeletons in 1996, which like PDW is out of print and I have no particular advice on where to obtain a copy. Sadly, the editor died at a young age.
Back in the 1980s I realized there was something wrong with Iguanodon, one of the iconic dinosaurs, being among the first two named back in the early 1800s to the fascination of the public. The problem started with the genus being based on a few non-diagnostic teeth. Then paleontologists chucked a wide array of remains from a broad span of time and around the northern hemisphere into Iguanodon. By the Reagan era three species based on good remains were among those still widely accepted; in Europe specimens tended to be split into the massive I. bernissartensis and lighter I. atherfieldensis, in America there was South Dakota’s I. lakotaensis.
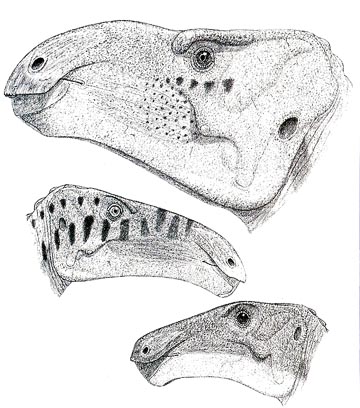 Now, I have long been concerned about how designating dinosaur genera has been and continues to be highly inconsistent. Many dinosaurs are oversplit at the genus level, while some others are over lumped, and there is little effort underway to deal with the problem. This was a classic case of putting very distinctive taxa with some striking anatomical differences in just one genus, and I planned to rename the Belgian I. bernissartensis. Did not get around to it. Let this be a lesson, procrastination can be a good thing. Unbeknownst to me the International Commission of Zoological Nomenclature had been petitioned to shift the type of the genus from Gideon Mantell’s teeth to the big Belgian beast. Whether this was a good idea or not was debatable (for one thing they are separated by about 10 million years in time and certainly do not represent the same genus), but had I renamed the robust iguanodont the ICZN ruling in favor of the petition would have invalidated the title. A few years ago I figured it was time to rename the gracile and American species before someone else got around to doing the obvious. While doing so, I realized I had been missing something. From the same Belgian quarry as robust species, the most complete specimen assigned to I. atherfieldensis, IRSNB 1551, had an ilium radically different in shape from the type specimen of the species over in England. And there were differences in the skull.
Now, I have long been concerned about how designating dinosaur genera has been and continues to be highly inconsistent. Many dinosaurs are oversplit at the genus level, while some others are over lumped, and there is little effort underway to deal with the problem. This was a classic case of putting very distinctive taxa with some striking anatomical differences in just one genus, and I planned to rename the Belgian I. bernissartensis. Did not get around to it. Let this be a lesson, procrastination can be a good thing. Unbeknownst to me the International Commission of Zoological Nomenclature had been petitioned to shift the type of the genus from Gideon Mantell’s teeth to the big Belgian beast. Whether this was a good idea or not was debatable (for one thing they are separated by about 10 million years in time and certainly do not represent the same genus), but had I renamed the robust iguanodont the ICZN ruling in favor of the petition would have invalidated the title. A few years ago I figured it was time to rename the gracile and American species before someone else got around to doing the obvious. While doing so, I realized I had been missing something. From the same Belgian quarry as robust species, the most complete specimen assigned to I. atherfieldensis, IRSNB 1551, had an ilium radically different in shape from the type specimen of the species over in England. And there were differences in the skull.
I did a dolt slap to my forehead - no one including me had restored the type I. atherfieldensis specimen on the assumption it was the same as the more complete IRSNB 1551. Duh. It was time to correct that data gap, and the restoration of the type specimen confirmed it was a different species from 1551, which would require a new name of its own. A pertinent example of the scientific utility of technical skeletal restorations. I published the results in Cretaceous Research, adding the names Mantellisaurus, Dollodon, and Dakotadon. As a result the once classic Brit dinosaur Iguanodon is barely known from the island.
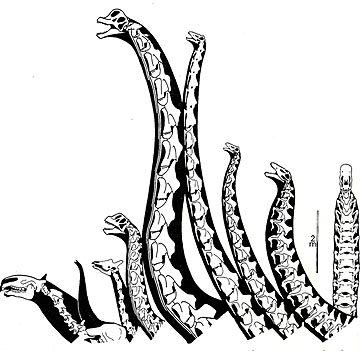 A controversy pertinent to both science and art became prominent in the late '90s and continues in the new century. Computer scientist Kent Stevens began to argue that sauropods habitually held their long necks horizontal, with the head often close to the ground. This is true of low-shouldered sauropods (contrary to the then radical Bakker illustration in 1968 showing a pair of striding barosaurs with their necks emerging nearly vertical from the horizontal dorsal columns), but they are a minority among the group. I can kick myself for not taking the opportunity to nip some of this stuff in the bud. When Stevens published his first Science paper I did not realize that some of the seemingly meticulous apatosaur cervical figures he used are actually fraudulent, Gilmore’s paper repeatedly states that the neck base vertebrae were badly crushed and embarrassingly incomplete. As I dealt with the issue I realized that the assumption we all shared that neck posture is accurately preserved in the articulations of the cervicals can be naive. True, the short necks of many theropods are so strongly S-curved that it has to reflect their basic posture, but long necks appear to be another matter. Some giraffe cervical series articulate highly erect, others subhorizontal, some with a peculiar downcurve. It appears to depend on the thickness of the intervertebral cartilage, which depends at least in part on the degree of ossification due to age. It is also being shown that long necked animals do not actually have a single neutral posture, giraffes hold their necks in whatever posture is suitable for the moment, so do ostriches. Because sauropods had so many neck vertebrae, flexing each pair just 10 degrees dorsally allows the long necked examples to hold their heads high over their backs. And the only fused sauropod cervicals, a pair from the base of the neck of an old Camarasaurus, are flexed upwards 10 degrees (on his website Stevens still contends these are articulated in a straight line). Turns out there is no simple set of rules by which artists can pose sauropod necks, it is more an optional thing. A paper in Anatomical Record will set this situation straight. Or is that more vertical?
A controversy pertinent to both science and art became prominent in the late '90s and continues in the new century. Computer scientist Kent Stevens began to argue that sauropods habitually held their long necks horizontal, with the head often close to the ground. This is true of low-shouldered sauropods (contrary to the then radical Bakker illustration in 1968 showing a pair of striding barosaurs with their necks emerging nearly vertical from the horizontal dorsal columns), but they are a minority among the group. I can kick myself for not taking the opportunity to nip some of this stuff in the bud. When Stevens published his first Science paper I did not realize that some of the seemingly meticulous apatosaur cervical figures he used are actually fraudulent, Gilmore’s paper repeatedly states that the neck base vertebrae were badly crushed and embarrassingly incomplete. As I dealt with the issue I realized that the assumption we all shared that neck posture is accurately preserved in the articulations of the cervicals can be naive. True, the short necks of many theropods are so strongly S-curved that it has to reflect their basic posture, but long necks appear to be another matter. Some giraffe cervical series articulate highly erect, others subhorizontal, some with a peculiar downcurve. It appears to depend on the thickness of the intervertebral cartilage, which depends at least in part on the degree of ossification due to age. It is also being shown that long necked animals do not actually have a single neutral posture, giraffes hold their necks in whatever posture is suitable for the moment, so do ostriches. Because sauropods had so many neck vertebrae, flexing each pair just 10 degrees dorsally allows the long necked examples to hold their heads high over their backs. And the only fused sauropod cervicals, a pair from the base of the neck of an old Camarasaurus, are flexed upwards 10 degrees (on his website Stevens still contends these are articulated in a straight line). Turns out there is no simple set of rules by which artists can pose sauropod necks, it is more an optional thing. A paper in Anatomical Record will set this situation straight. Or is that more vertical?
In 2000 I connected with Hall Train, who has a studio near Toronto. We have collaborated on a number of projects, the largest being the large dinosaur hall installed in the new wing of the Maryland Science Center (MSC) completed in 2004. Among other items I designed the full-sized sculptures of Acrocanthosaurus attacking Astrodon in the famous Paluxy trackway. On December 5th 2002 I was scheduled to fly with a person from the MSC to Miami to inspect part of a traveling dinosaur show they were thinking of purchasing for the new hall. An unusually early snowstorm hit Baltimore and made getting to and from the airport something of an adventure. Exactly a year later by coincidence a MSC team including myself drove to the Philly suburbs to inspect the purchase before delivery. Again a major snowstorm hit the region making it a winter adventure.
 During this period the MSC decided to issue a special Maryland license plate with my illustration of the state dinosaur, Astrodon, on it. I assume I am the only dinosaurologist whose car has a license plate bearing an image he made on it. Have worked with Animatronix in Mexico City on full scale dinorobots. Did I ever tell you about the time fellow consultant Jim Kirkland got sick as a dog in MSC, poor fellow?
During this period the MSC decided to issue a special Maryland license plate with my illustration of the state dinosaur, Astrodon, on it. I assume I am the only dinosaurologist whose car has a license plate bearing an image he made on it. Have worked with Animatronix in Mexico City on full scale dinorobots. Did I ever tell you about the time fellow consultant Jim Kirkland got sick as a dog in MSC, poor fellow?
In 2007 Ian Paulsen complained on the Dinosaur Mailing List about the lack of a high quality dinosaur field guide. I agreed. That led to contact with Princeton University Press and a contract to author and illustrate such a volume. This is an excuse to complete as many skeletal restorations as is feasible, getting me my closer to the long standing goal. Doing so is often vexing because many species known from good specimens have yet to be sufficiently documented. Take Majungasaurus, which is known from almost perfectly preserved uncrushed remains. But none of the specimens is complete enough to entirely scale out its proportions, and a new more complete specimen has yet to be prepped out. Some of the feathered Yixian dinosaurs are known from complete but badly crushed specimens that lack detail.
One thing I have been doing is being specific to the specific age and level a particular species of dinosaur comes from. For example, Allosaurus fragilis has long been assigned to most or all of the formation. But radiometric dating has shown that the formation formed over a span of 7 million years or more, and species rarely last that long. And the Allosaurus skulls show tremendous variation, from short and stout as in A. fragilis to very long, low and elegant. A. fragilis is therefore limited to the lower Morrison.
Creature Features - in Shocking 2-D
Because a guide emphasizes anatomical features that can be used to identify a given species, it has been necessary to not do the kind of dramatic color features on the color pencil studies I often apply to other color works lest I mislead the reader into thinking the color patterns are real. The latter is a real danger. The question paleoartists most often receive is how do we know what color the fossil animal was. This is, of course, illogical. If we knew what color a given extinct species was then all artists would color them the same way.
In my view the most frustrating thing about paleontological science and art is that because of the way our particular universe is constructed there is no practical way of going back in time and seeing what dinosaurs actually looked liked and behaved, and what their habitats were really like. On the other hand traveling back to the Tendaguru, Morrison or Hell Creek as a human being would have its downsides, even assuming issues of novel diseases could be dealt with. The great dinosaurs would be so dangerous that encampments would probably have to be fortified, and moving about the landscape would require being ensconced in reinforced vehicles. There is also the possibility that oxygen levels were low compared to current sea level standards well into the Cretaceous.
Dinosaurology is many, many times more expansive and active than it used to be, and a lot of good work is being done as the methods generally improve. I say generally, because there are problems. Cladistics is very useful, but characteristic counts cannot determine actual relationships beyond a certain level for a number of inherent reasons, and the methods are being applied too rigidly. At the other extreme the continued attempts to separate birds from dinosaurs uses pseudo-logic that borders on Creationism. Bone rings are being used to assign ages to dinosaur specimens even though the technical literature warns against doing so without field observations because the two do not match in all birds. Computers are powerful tools, but they can seduce those who use them into accepting dubious results. I have seen computer generated technical simulations of sauropod locomotion that show them lurching about with flexed elbows like ungulates rather than the straight, elephantine limbs they clearly had. Sometimes a little common sense is called for.
At the other side of the limb flexion issue is that some continue to show all big dinosaurs with straight legs on the premise they were de rigueur to support the animal’s mass. But the anatomy of dinosaur joints shows that only sauropods and some stegosaurs had elephantine legs, in all others the elbows and knees were more flexed. I continue to roll my eyes at many new and revised skeletal mounts, which often violate basics of tetrapod anatomy, or well established facets of dinosaur form and function. Many who construct dinosaur mounts do not have a scientific understanding of form and function. Some brand new mounts even have the sternals on backwards despite mind the numerous technical papers (mine among them) showing the correct placement.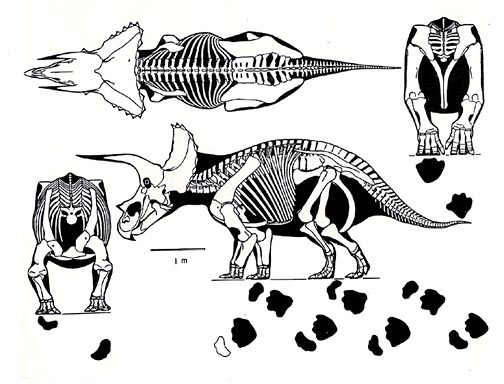
At the other extreme some take the New Look too far, showing giant ceratopsids bounding about like gazelles, and sauropods trotting or pacing like horses.
(Note to artists: Never, ever, apply wrinkled elephant skin to sauropods, which were not elephants.)
The failure to place the feet in accord with the position and posture recorded in trackways is most vexing. Gaits are often of an arrangement not seen in real animals, and the four feet all on the ground is still a problem. No erect-limbed animal has all four feet on the ground at any time when moving.
The output of dinosaur art is many multiples of what it used to be. Because I go to such great technical efforts to determine dinosaur anatomy and function I can be critical of the accuracy of other artist’s work, the artistic quality of which sometimes inspires my admiration, and other times does not. Among my favorite contemporary paleoart are the B&W fossil mammals by Mauricio Anton, which channel those of Matternes. A particular pet peeve is how small dinosaurs are often illustrated with exposed eyes filling almost the entire orbit.
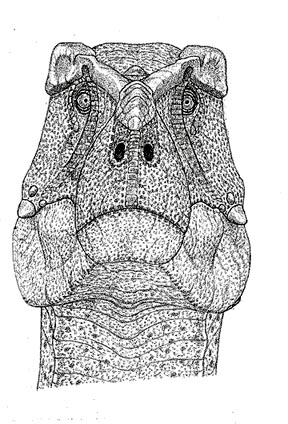 That only occurs among a limited number of very small reptiles, in dinosaurs eyelids should have covered most of the eyeball, which had limited mobility like in birds. Nor do I favor exposed crocodile type teeth in theropods and sauropods, they were probably covered by lips as in tuataras, lizards and snakes.
That only occurs among a limited number of very small reptiles, in dinosaurs eyelids should have covered most of the eyeball, which had limited mobility like in birds. Nor do I favor exposed crocodile type teeth in theropods and sauropods, they were probably covered by lips as in tuataras, lizards and snakes.
Paleoflora is often a problem too as paleobotanists have yet to present enough information in a comprehensive format, although they note that the umbrella-shaped monkey puzzle conifers is overdone. I have developed a corresponding penchant for cloud-filled skies.
Alas, producers of commercial dinosaur products continue to churn out low quality product that is either obsolete or improperly derivative. Dino documentaries and books have become so plentiful that they are no longer special and I do not try to keep up with them. There are also serious problems with quality and accuracy which often fail to meet the expectations of scientists. More about those problems here. I about kicked in the TV screen when one dino doc claimed that the brain of Tyrannosaurus was as large as that of a gorilla when its IQ was not all that much better than a croc’s. And why are the theropods shown pausing to challenge their prey before they charge, when the actual focus of predators is to hit and overwhelm the victim before it knows what is happening? The low standards are not surprising considering how the media and press frequently carry product that promotes belief in the paranormal.
But these are quibbles. Dinosaur science has almost completely transformed over the half century that my neural network has been aware of it. The old stand-bys from Allosaurus to the always strange Stegosaurus are still fascinating, but we now know about armored sauropods, fat-bellied therizinosaurs and multi-winged, near avian, sickle claws. The reptile model is out and the avian-mammalian is dominant.
It is a fascinating scientific and artistic ride.
 Gregory Scott Paul
Gregory Scott Paul
Baltimore, Md.
June 2009
